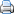|
ALTRES FITXES DE "La Planta del Mes" PUBLICADES AL PORTAL BIOC
Esperó de muntanya
Per Maria Bosch
Delphinium montanum DC. in Lam. & DC., Fl. Fr. 5: 641 (1815)

Individus de Delphinium montanum (Fotos: esquerra, M. Bosch i dreta, J. López-Pujol)
L'esperó de muntanya (Delphinium montanum, Ranunculaceae) és una espècie perenne, tetraploide, endèmica dels Pirineus orientals, restringida a 9 poblacions (3 de les quals no s’han tornat a trobar recentment) i amenaçada (vulnerable [VU] segons criteris de la UICN de 2001). Tàxon objecte d’estudi per l’equip del BioC des de fa uns 25 anys, des del punt de vista taxonòmic (Blanché, 1991), de biologia reproductiva i diversitat genètica (Bosch, 1999; Simon et al., 2001) i de l’origen de la tetraploïdia (López-Pujol et al., 2007). Ha format part de diversos projectes de recerca entre els quals destaca “Conservación de la biodiversidad en zonas sometidas a impacto del sector turístico y fragmentación de poblaciones. Bases para la gestión de especies amenazadas”, finançat pel Plan Nacional de I+D del Ministeri d'Educació i Ciència.
Descripció
Herba perenne, rizomatosa, amb una soca subterrània desenvolupada. Tiges de (15) 30-50 (90) cm. Les fulles simples, palmades i laciniades formen una roseta basal de la qual, en l’època de floració, es produeixen 1 (2-3) raïms de (3-5) 15-20 (50) flors. De vegades, alguns individus formen rodals compactes amb nombroses tiges floríferes. Les flors, hermafrodites i marcadament zigomorfes, presenten un esperó posterior nectarífer de 11-15 cm. El periant és petaloide de color blau cel (o en ocasions blanc), amb 4 pètals centrals (2 de nectarífers, interns, i 2 de laterals, externs, de color bru fosc i amb un indument ciliar blanc-groguenc). La coloració fosca d’aquestes peces és una particularitat que només es presenta en el subgènere Delphinastrum i que, junt amb la pilositat blanc-groguenca, serveixen de guies de nèctar, recordant la morfologia d’un àpid (síndrome de la bee-flower). El fruit presenta 3 fol·licles densament pubescents que produeixen llavors petites negres subpiramidals, amb petites expansions alades a les arestes.
• Nombre cromosòmic: tetraploide amb 2n = 32 (Blanché, 1991)
• Fenologia: floreix entre juliol i agost

Detall de la inflorescència i de les flors de Delphinium montanum (Fotos: M. Bosch)
Taxonomia
Delphinium montanum és una espècie de la família de les Ranunculàcies, endèmica dels Pirineus Orientals, que pertany a la sèrie Montana B. Pawl del subgènere Delphinastrum (DC) Peterm. Així doncs, taxonòmicament està emparentat amb D. dubium B. Pawl. i amb D. oxysepalum Borb. & Pax, endemismes dels Alps i els Càrpats, respectivament. S’ha proposat que les tres espècies provenen d’un avantpassat comú (molt proper a D. elatum) que hauria donat lloc als tres taxons per fragmentació de l’àrea abans de la darrera glaciació (Pawlovsky, 1970). Totes aquestes espècies són tetraploides amb 2n = 32 cromosomes.
Sinònims del nom científic:
≡ Delphinium elatum subsp. montanum (DC.) Nyman
≡ Delphinium elatum var. montanum (DC.) Rouy & Fouc.
- Delphinium elatum auct.
- Delphinium pyrenaicum Pourr. (nom. inval.)
Distribució geogràfica
L’esperó de muntanya és un endemisme dels Pirineus orientals, principalment de la vora S. i E. de la fossa de la Cerdanya (serra del Cadí i massís del Puigmal), amb extensions fins al Pedraforca i el massís de Madres, cobrint una àrea de 60 km de llargada i uns 15 d’amplada, en territori administrativament espanyol i francès. Es coneix de nou localitats (tres de no retrobades recentment). A la Catalunya autònoma té presència confirmada actual en tres localitats: dues de la serra del Cadí (capçalera de la vall de Bastanist (Cerdanya) i serra Pedregosa (Berguedà)) i una del massís del Puigmal (Coma de l’Orri, vall de Setcases, Ripollès). S’havia observat en una altra localitat del Cadí (Cava, Alt Urgell) no retrobada des del 1985, però insuficientment prospectada i al Pedraforca (Berguedà), població no retrobada des de 1984 i probablement extingida. A la Catalunya del Nord es troba en dues localitats a la serra del Puigmal (vall d’Eina i Cambra d’Ase), una al massís de Madres (Noedes) i una al coll de la Portella de Mantet, no retrobada des de 1896. Les indicacions antigues del massís de Sant Llorenç del Munt, provinents d’una citació de Cadevall de principis del s. XX, són errònies i corresponen en realitat a D. bolosii (Blanché, 1991).
Mapa de les localitats de Delphinium montanum : 1) Bastanist, 2) Els Cortils, 3-4) Vall d’Eina, 5) Coma de l’Orri i 6) Noedes (Font J. López-Pujol)

Imatges de les localitats de Delphinium montanum. Dreta:1) Bastanist; Esquerra: 6) Noedes. (Fotos: J. López-Pujol)

Imatges de les localitats de Delphinium montanum. Dreta: 2) Els Cortils; Esquerra: 5) Coma de l'Orri. (Fotos: J. López-Pujol)

Imatges de les localitats de Delphinium montanum. 3-4) Vall d'Eina (Fotos: J. López-Pujol)
Hàbitat
Pel que fa a l’ecologia es troba en espais rocosos oberts, entre 1600 i 2200 (2400) m, sobre substrat calcari mòbil, principalment tarteres més o menys estabilitzades (Aquilegio-Xatardietum scabrae) i peus de cingle (Ranunculion thorae). Pot penetrar, en tarteres fixades, a comunitats megafòrbiques (Delphinieto-Trollietum, Adenostylion alliariae). Pot tolerar graus moderats de nitrificació.

Hàbitat de Delphinium montanum: De dalt a baix, a i b) Serra del Cadí, c) Vall d’Eina (Fotos: M. Bosch)
Pol·linització, reproducció i estratègia vital
Pel que fa als pol·linitzadors, es va realitzar un estudi observacional de 84 períodes diferents de 15 minuts cadascun en dues localitats (Els Cortils, a la serra del Cadí i a la Vall d’Eina) (Bosch, 1999). Es van censar 411 visites. Els pol·linitzadors més freqüents i més efectius van ser els abellots de l’espècie Bombus hortorum hortorum (50 % de les visites), gràcies a la seva llarga probòscide, que els permet assolir el nèctar amagat a l’esperó, i al seu cos robust ,que els permet separar les peces florals i empolsinar-se del pol·len situat sota dels pètals laterals. En canvi, Bombus wurfleini pyrenaicus (9% de les visites), atès que té una probòscide més curta que li limita accedir al nèctar de l’esperó, ha après a foradar externament aquesta peça floral; per tant, actua com a un robador de nèctar, ja que no visita la flor per la via legítima i d’aquesta manera no contacta amb els estams o ho fa de forma més accidental. Tots els exemplars observats van tenir el mateix comportament. Altres pol·linitzadors menys efectius són dípters (diferents espècies de sírfids) (35% de les visites), algunes petites abelles solitàries del gènere Lassioglossum (3,6%), o lepidòpters com l’esfíngid Macroglossum stellatarum (1,8%).

Pol·linitzadors de Delphinium montanum: 1) Bombus hortorum hortorum

Pol·linitzadors de Delphinium montanum: 2) Bombus wurfleini pyrenaicus

Pol·linitzadors de Delphinium montanum: 3) Macroglossum stellatarum 4) sírfid (Fotos: M. Bosch)
Recompenses florals: nèctar i pol·len
Per tal d’atraure els insectes pol·linitzadors, l’esperó de muntanya és capaç de produir recompenses suficients per a la dieta dels insectes visitants. S’ha detectat una producció mitjana de nèctar de 0,9 ± 0,5 µl (m ± SD) per dia i una concentració de sucres, estimada per refractometria, lleugerament superior al 45% (v/v), amb una disminució del volum al llarg de la inflorescència (Bosch, 1999). La flor presenta entre 20-24 estams que produeixen al voltant de 86.000 grans de pol·len per flor.

Mesura del nèctar contingut als esperons per mitjà de micropipetes (Foto: M. Bosch)
Relació pol·len / òvul (P/O)
Aquest paràmetre té un valor mitjà de 2.125 a D. montanum, la qual cosa ens indica, segons Cruden (1977, Evolution 31: 32-46), un sistema reproductiu al·lògam facultatiu. Els resultats de l’estudi del sistemes reproductius, mitjançant l’embossament d’inflorescències en les poblacions naturals, mostren una producció de llavors pràcticament nul·la, indicant una important dependència dels pol·linitzadors per a la seva reproducció (Bosch, 1999; Bosch et al., 2001).
Estudis de germinació
La viabilitat (teòrica) de les granes produïdes per l’esperó de muntanya ha estat estimada com a molt alta (90-92%) per Aymerich (2003), en base al test de tetrazoli. No obstant això, els estudis de germinació que es van dur a terme en germinadors en condicions controlades d’hivernacle, sense cap tractament previ a les llavors, van donar uns resultats molt minsos, al voltant d’un 5-6% (N = 216 llavors, Bosch, 1999). Es van realitzar proves de germinació en germinadors ubicats les poblacions naturals i els resultats van ser lleugerament superiors, 6-12% (N = 360 llavors). La germinació triga uns 145 dies de mitjana en tenir lloc. Les llavors presenten una dormició de quasi tres mesos, que coincideix amb el període en què les poblacions solen estar cobertes de neu i no es pot descartar que sigui una estratègia de retardar la germinació fins al moment adequat i assegurar una probabilitat superior de supervivència de les plàntules; a més, la biologia del desenvolupament de l’embrió indica que, en el moment de la disseminació, no ha finalitzat el procés de maduració, que s’acabarà en una etapa de post-dispersió, com en d’altres ranunculàcies (cf. Blanché, 1991).

Estudis de germinació en les poblacions naturals (Foto: M. Bosch)
Malgrat la forta predació de les tiges floríferes per part dels isards, es produeix reclutament i s’observen plàntules cada any en les poblacions naturals.

Plàntules de Delphinium montanum (Foto: M. Bosch)
Les prospeccions del banc de granes del sòl dutes a terme per Aymerich (2003) a les poblacions del Cadí van donar resultats negatius, de manera que cal assumir que les noves plàntules han de provenir de la producció de l’any immediatament anterior, originades de les flors i fruits que han escapat de la predació (l’absència de granes al banc de sòl podria ser motivada també per l’exhauriment de reserves en no haver-hi prou incorporació de noves granes a causa de la depredació -vegeu més endavant- atès que els resultats de germinació al camp en condicions naturals indiquen que més d’un 80% de les granes resten al sòl sense emergir).
Segons Aymerich (l.c.), la majoria (prop de 80%) de les granes que germinen al camp ho fan al final del període vegetatiu (finals de juny) i la resta germinen esglaonadament fins a l’agost, aconseguint densitats de (1) 2,5 (5) plàntules/m2 i de (2) 7,2 (13) plàntules/m2 en dues poblacions estudiades; la mortalitat de les plàntules a finals de setembre és del 52-65% de mitjana a escala de població (variable entre parcel·les: 0-100%).
Diversitat genètica
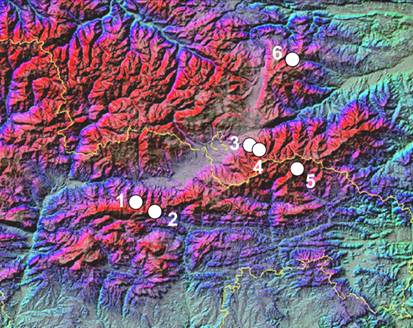
S’han estudiat els nivells i la distribució la diversitat genètica d’aquesta espècie tetraploide mitjançant l’electroforesi d’isoenzims (Simon et al., 2001; López-Pujol et al., 2007). S’han mostrejat 7 poblacions i s’han resolt 8 enzims (AAT, ACP, DIA, IDH, MDH, 6PGD, PGI i PGM) que han proporcionat 14 loci interpretables (Aat-1, Aat-2, Acp-2, Dia, Idh-1, Mdh-1, Mdh-2, 6Pgd-1, 6Pgd-2, 6Pgd-3, 6Pgd-4, Pgi-2, Pgm-1 i Pgm-2).
Els patrons de bandes obtinguts presenten moltes duplicacions atribuïbles a un origen autotetraploide d’aquesta espècie, també refermat per la presència d’heterozigots equilibrats i desequilibrats i per l’absència de fixació de l’heterozigosi. No obstant això, la variabilitat de D. montanum (P = 24,5%, A = 1,49, i He = 0,088) és més baixa que l’esperada en espècies autotetraploides. La distribució de la diversitat és l’esperada per a espècies amb poblacions genèticament aïllades (fins un 15% de la diversitat es deu a diferències entre poblacions). La baixa diversitat observada, juntament amb la baixa freqüència de loci que presenten 3 o 4 al·lels diferents pot indicar que aquesta espècie ha sofert erosió genètica a través de colls d’ampolla o, alternativament, que s’ha produït certa diploidització. Altres indicis, com ara la morfologia del cariotip (les parelles cromosòmiques són una mica diferents), la no observació de tetravalents a la meiosi o d’altres característiques morfològiques (mida del gra de pol·len, mida dels estomes i nombre d’estams) suggereixen aquesta diploidització (Blanché, 1991).
Patrons de bandes obtinguts per l’Aat-1. Es poden observar tant homozigots com heterozigots equilibrats i desequilibrats. (Font: López-Pujol et al., 2007)
|
Localitat
|
P
|
A
|
Ho
|
He
|
F
|
|
BAS
|
21,4
|
1,36
|
0,050 (0,092)
|
0,065 (0,122)
|
0,232 (0,075)
|
|
COR
|
21,4
|
1,64
|
0,076 (0,130)
|
0,102 (0,176)
|
0,253 (0,094)
|
|
ORR
|
28,6
|
1,50
|
0,104 (0,191)
|
0,098 (0,162)
|
-0,056 (0,063)
|
|
EI1
|
28,6
|
1,43
|
0,071 (0,140)
|
0,073 (0,142)
|
0,027 (0,013)
|
|
EI2
|
28,6
|
1,50
|
0,083 (0,140)
|
0,095 (0,145)
|
0,121 (0,074)
|
|
CAS
|
21,4
|
1,57
|
0,095 (0,201)
|
0,107 (0,199)
|
0,107 (0,135)
|
|
NOE
|
21,4
|
1,43
|
0,081 (0,175)
|
0,074 (0,142)
|
-0,087 (0,068)
|
|
Mitjana
|
24,5
|
1,49
|
0,080
|
0,088
|
0,085
|
|
Desviació estàndard
|
3,8
|
0,09
|
0,017
|
0,017
|
0,132
|
Resum de la diversitat genètica de 14 loci en set poblacions de Delphinium montanum.
Codis: BAS, Bastanist; CAS, Cambra d’Ase; COR, Els Cortils; EI1, Vall d’Eina (Inferior); EI2, Vall d’Eina (Superior); NOE, Noedes; ORR, Coma de l’Orri. P, percentatge de loci polimòrfics; A, nombre mitjà d’al·lels per locus; Ho, heterozigosi observada; He, heterozigosi panmíctica esperada; F, mitjana del índex de fixació. Desviació estàndard en parèntesi (Font: López-Pujol et al., 2007).
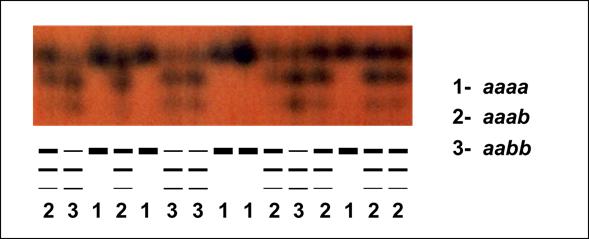
El dendrograma UPGMA de les distàncies genètiques de Nei ens separa clarament la població de Noedes de la resta. Això es pot deure a que aquesta població ha estat sotmesa a un aïllament genètic més gran. Si hom observa el mapa de relleu, mentre que totes les poblacions estan situades en l’eix muntanyós format per la serra del Cadí i el massís del Puigmal, la població de Noedes està localitzada en el massís de Madres, separada manifestament de l’anterior eix per la fossa de Prades. Les altres poblacions més al sud, per tant, han tingut majors probabilitats d’intercanvi genètic, mentre que la població de Noedes pot haver quedat aïllada genèticament de la resta com a mínim des de la darrera glaciació i haver evolucionat de forma independent. Aquesta singularitat recomana la conservació diferenciada de la població de Noedes, en particular en les iniciatives de recol·lecció de germoplasma.
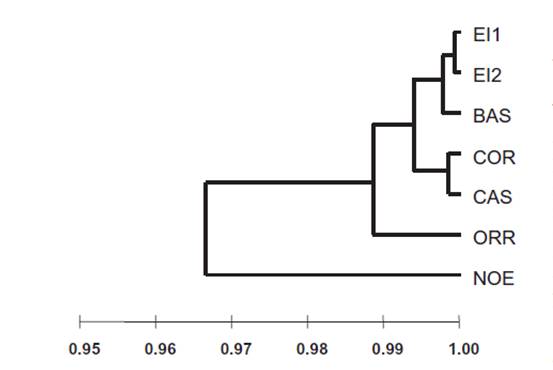
Dendrograma resultant de l’anàlisi UPGMA de distàncies genètiques de Nei per les poblacions de D. montanum (Font: López-Pujol et al., 2007)
Demografia
L’estructura demogràfica de les poblacions ha estat estudiada a instàncies del Parc Natural del Cadí-Moixeró (Aymerich, 2003) i, en resum, hom pot concloure que es tracta de poblacions constituïdes principalment per individus adults (depenent de les poblacions: juvenils= 2,7-20,7%; no reproductors= 21,4-22,9%; adults reproductors= 56,4-75,9%). Aquest autor ha observat una certa heterogeneïtat entre nuclis dins una mateixa població i destaca que les menors taxes de regeneració (% d’adults >> % juvenils) es donen en indrets de tarteres poc o gens mòbils, amb blocs de pedra fixats o afloraments rocosos estables, mentre que els fragments més mòbils, pertorbats i amb vegetació poc densa afavoreixen la instauració de noves plàntules, suggerint que la disponibilitats d’hàbitat adequat a la germinació serien el factor clau en el reclutament.
Predació
L’esperó de muntanya és una planta sotmesa a una predació notable de flors i fulles (especialment les tiges florides) per part fonamentalment d’isards (Rupicapra pyrenaica), que pot fluctuar molt d’una localitat a un altra o d’un any a un altre segons la disponibilitat d’altres pastures a l’alta muntanya pirinenca. La densitat d’isards hi ha arribat a ser molt elevada durant els mesos d’estiu, amb densitats habituals de 40-60 animals per km2 en la zona immediata a la població de D. montanum i amb màxims ocasionals de fins a 100 per km2 (Aymerich, 2003); els percentatges d’afectació poden arribar a superar el 90% de les flors d’una població en un any determinat (Simon et al., 2001).
Cal tenir en compte que es tracta d’una planta que conté alcaloides diterpènics (amb toxicitat similar a la dels acònits, una de les plantes més tòxiques de la flora europea) i que podria ser causant de la mort d’aquests animals per ingestió, atès que s’han trobat alguns cadàvers enmig de les poblacions d’esperó de muntanya. No obstant això, caldria confirmar la relació causa-efecte de manera concloent, atès que les parts tendres solen contenir menys alcaloides i menor activitat tòxica (Olsen et al., 1990) i que hi ha d’altres causes de mortalitat d’aquests animals.

Dreta: individu depredat de Delphinium montanum (hi manquen les flors superiors de la inflorescència) Esquerra: isard mort en una localitat d'esperó de muntanya.
A partir d’aquestes dades, havíem suggerit (Simon et al., 2001) que al Parc natural del Cadí-Moixeró podia existir un conflicte de gestió entre els isards i Delphinium montanum, que, en canvi, no s’observava a la Reserva natural de la Vall d’Eina (vaques i cavalls eviten la ingestió de brots de l’esperó de muntanya). A causa d’aquesta preocupació, els responsables del PN del Cadí van encarregar els informes científico-tècnics pertinents que van ser donats a conèixer per Aymerich (2003) en un treball on estudia l’impacte de la depredació de dos ungulats (Rupicapra pyrenaica Bonaparte i Ovis ammon L.) sobre l’esperó de muntanya. L’estudi confirma que en sectors amb alta densitat d’ungulats la incidència de la depredació és forta i afecta molt especialment les estructures reproductores de les plantes (en un cas extrem, la pèrdua de fruits va ser estimada en un 98,9 % de la producció per a una població de D. montanum).
Segons Aymerich (l.c.), tanmateix, no sembla que aquest consum afecti el reclutament ni la viabilitat de les poblacions. La comparació entre les estructures de poblacions amb diversa incidència dels ungulats (molt depredades i poc depredades) indica que el nombre de plantes juvenils no està relacionat amb el nombre de granes produïdes i que un factor determinant en el percentatge de plàntules sembla la disponibilitat de substrats adequats per a germinar, de manera que la mortalitat de les plantes joves –probablement per competència intraespecífica– conduiria finalment a unes estructures poblacionals adultes. No obstant això, hom no pot descartar que el consum del possible estoc de granes del banc del sòl que (malgrat que produeixi individus florífers ,acabi essent depredat i no acabi contribuint a la generació següent) afecti a llarg termini els nivells de diversitat genètica (per limitació del nombre d’encreuaments disponibles entre genotips diferents). Aquest fenomen, però, es veuria diferit en el temps a causa de la longevitat dels individus i de l’efecte tampó de la poliploïdia (López-Pujol et al., 2007). Observacions posteriors confirmen que l’espectacular depredació es dóna també a les poblacions de Cambra d’Ase i de Noedes (pràcticament totes les tiges depredades) mentre que Oliver (2008) reporta pocs efectes a la població del Ripollès. En tot cas, caldria reavaluar les conseqüències a llarg termini d’aquest fenomen de depredació i el Parc Natural del Cadí-Moixeró ha emprès, des d’aleshores, mesures per a reduir la densitat i la pressió dels isards sobre l’esperó de muntanya.
A més de l’afectació de les inflorescències, també hi ha un consum important de fulles, exercit tant pels ungulats com pel rosegador Chionomys nivalis sobre D. montanum (Aymerich, l.c). Les llavors també sofreixen una predació important per erugues de lepidòpters (11-42%, Aymerich, l.c.) i per larves de dípters, que les barrinen i les malmeten notablement (Bosch, 1999).
Llavors de l’esperó de muntanya barrinades (Foto: M. Bosch)
Conservació
Població
La darrera avaluació (Sàez et al., 2010) de l’estat de conservació de l’esperó de muntanya indica presència confirmada actual a la Catalunya autònoma en només 3 quadrats UTM de 10 x 10 km, que inclouen uns 5.000 individus adults reproductors en diverses localitats de la serra del Cadí, de la capçalera del Ter, del massís del Pedraforca i de la vall de Camprodon; a la Catalunya del Nord s’hi compten uns altres 4.000 individus (serra del Puigmal, massís de Madres, Cambra d’Ase i vall d’Eina). L’extensió de presència és de 827,5 km2 i l’àrea d’ocupació total <20 km2 (López-Pujol et al., 2007).
Amenaces
- Pel que fa als factors biòtics, s’han constatat oscil·lacions de la mida poblacional (variacions de cens per desaparició temporal i reaparició, al cap d’un temps, en part lligada a la mobilitat del substrat); nuclis poblacionals envellits probablement per la dinàmica contrària (d’estabilització de tarteres).
- Existeix també risc d’afecció per sobrefreqüentació de visitants a les poblacions situades en indrets d’interès natural a l’estiu (trepig, recol·lecció i sobretot degradació de l’hàbitat, particularment notòria a la població de la canal del Pedraforca, on no ha estat retrobada des de 1984). S’han descrit episodis de predació de fulles i flors per isards (sobrepastura) als Parcs Naturals del Cadí-Moixeró i Noedes (Simon et al., 2001; López-Pujol et al., 2007), tot i que aquesta afectació ha estat avaluada com a poc rellevant per Aymerich & Sàez (2001) i Aymerich (2003), confirmada per dades de 2007 (Aymerich, com. pers.). i amb escàs impacte (Oliver, 2008).
Avaluació del risc d’extinció
L’espècie ha estat avaluada en diverses ocasions per a les poblacions de la Catalunya autònoma. Considerada "Vulnerable" (VU) B1+2e, C1 (criteris UICN del 1994) en una avaluació prèvia de la flora endèmica de Catalunya (Sáez et al., 1998) i a la Lista Roja de la Flora Vascular Española (Aizpuru et al., 2000), i "Vulnerable" (VU) B2ac(iii), D2 (criteris UICN 2001) en la darrera Lista Roja del 2008 (Moreno, 2008). L’última avaluació disponible, en el Llibre Vermell de Catalunya, la qualifica com a "Vulnerable" (VU) B2ac(iii), C1, D2 (Sáez et al., 2010). També és considerada, pel que fa a les poblacions de la Catalunya del Nord, com a "Vulnerable" (VU) en el segon volum del Livre rouge de la flore menacée de France (Olivier et al., 1995).
Protecció legal
- Les poblacions de la Catalunya autònoma gaudeixen de protecció legal en tractar-se d’una espècie inclosa al Catàleg de Flora Amenaçada de Catalunya (DOGC, 2008), en la categoria de “Vulnerable” (Annex 2)
- Totes les poblacions (també les no retrobades) es troben en espais natural protegits: Parc Natural del Cadí-Moixeró o espais PEIN (a la Catalunya autònoma), Reserves Naturals d’Eina, Noedes, etc. (a la Catalunya del Nord).
Mesures de conservació
Accions realitzades
• Seguiment i censos per part d’investigadors del BioC
• Estudis de l’efecte de la predació i d’estructura demogràfica de poblacions per part del Parc natural del Cadí-Moixeró
• Regulació de la densitat d’isards per part del Parc natural del Cadí-Moixeró
• Seguiment de les poblacions del Ripollès per part de la Delegació de la Garrotxa de la ICHN
• Llavors conservades en els bancs de germoplasma de la UB i del Jardí Botànic Marimurtra
Mesures proposades
• Redacció del Pla de Conservació de l’espècie derivat de la inclusió al Catàleg de Flora Amenaçada (DOGC, 2008)
• Continuació del seguiment poblacional i avaluació periòdica de l’efecte de la predació per part dels isards
• Regulació de l’accés a certes poblacions
• Coordinació de les activitats de conservació de les diverses administracions implicades (dos estats de la UE, dues administracions regionals, cinc espais protegits) prioritzant les ESU identificades (López-Pujol et al., 2007)
• Conservació de llavors en bancs de germoplasma amb representativitat de la diversitat genètica detectada (noves accessions)
• Conreu en jardins botànics alpins
Bibliografia
Aizpuru, I. et al. 2000. Lista roja de la flora vascular española (valoración según categorías UICN). Conservación Vegetal 6 (extra): 11-38.
Aymerich, P. 2003. Efectes de la depredació dels ungulats salvatges en la conservació de plantes rares: dos casos dels Pirineus orientals. Acta Botanica Barcinonensia 49: 147–164.
Aymerich, P. & L. Sáez. 2001. Dades sobre l’estatus d’algunes plantes endèmiques, amenaçades o rares a Catalunya (NE de la península Ibèrica). Orsis 16: 47–70.
Blanché, C. 1991. Revisió biosistemàtica del gènere Delphinium L., a la Península Ibèrica i a les Illes Balears. Arxius de la Secció de Ciències 98, Institut d’Estudis Catalans, Barcelona.
Bosch, M. 1999. Biologia de la reproducció de la tribu Delphinieae a la Mediterrània occidental. Arxius de la Secció de Ciències 120, Institut d’Estudis Catalans, Barcelona.
Bosch, M., J. Simon, C. Blanché & J. Molero. 2001. Breeding systems in the tribe Delphinieae (Ranunculaceae) in the Western Mediterranean area. Flora 196: 101-113.
DOGC. 2008. Decret 172/2008, de 26 d’agost, de creació del Catàleg de flora amenaçada de Catalunya. DOGC Núm. 5204-28.8.2008.
López-Pujol, J., M.R. Orellana, M. Bosch, J. Simon & C. Blanché. 2007. Low genetic diversity and allozymic evidence for autopolyploidy in the tetraploid Pyrenean endemic larkspur Delphinium montanum (Ranunculaceae). Botanical Journal of the Linnaean Society 155: 211-222.
Moreno, J. (coord.). 2008. Lista Roja 2008 de la flora vascular española. Dirección General de Medio Natural y Política forestal (Ministerio de Medio Ambiente y Medio Rural y Marino) y Sociedad Española de Biología de la Conservación de Plantas. Madrid, 86 pàgs.
Oliver, X. 2008. Delphinium montanum, fitxa de seguiment. Programa de seguiment i conservació de la flora amenaçada de la Garrotxa. Delegació de la Garrotxa de la Institució Catalana d’Història Natural. Inèdit.
Olivier, L., J.P. Galland & H. Maurin. 1995. Livre rouge de la flore menacée de France, Tome I. Ministère de l’Environnement. Paris.
Olsen, J.D., G.D. Manners & S.W. Pelletier. 1990 Poisonous Properties of Larkspur (Delphinium spp.). Collectanea Botanica. (Barcelona) 19: 141-151.
Pawlovski, B. 1970. Remarques sur l’endemisme dans la flore des Alpes et des Carpates. Vegetatio 21: 181-243.
Sáez, L., P. Aymerich & C. Blanché 2010. Llibre Vermell de les plantes vasculars endèmiques i amenaçades de Catalunya. Argania editio, Barcelona. 811 pàgs.
Sáez, Ll., J. A. Rosselló & J. Vigo. 1998. Catàleg de plantes vasculars endèmiques, rares o amenaçades de Catalunya, I. Tàxons endèmics. Acta Botanica Barcinonensia 45: 309-321.
Simon, J., M. Bosch, J. Molero & C. Blanché. 2001. Conservation biology of the Pyrenean larkspur (Delphinium montanum): a case of conflict of plant versus animal conservation? Biological Conservation 98: 305–314.
ACTUALITZACIÓ 2/10/2012
D. montanum ha estat retrobat al Massís del Pedraforca per J. Torra (un únic individu l'estiu de 2009)
Més informació
CITACIÓ / PDF / HANDLE
ALTRES FITXES DE "La Planta del Mes" PUBLICADES AL PORTAL BIOC
Taxonomia
Delphinium montanum és una espècie de la família de les Ranunculàcies, endèmica dels Pirineus Orientals que pertany a la sèrie Montana B. Pawl del subgènere Delphinastrum (DC) Peterm. Així doncs, taxonòmicament està emparentat amb D. dubium B. Pawl. i D. oxysepalum Borb. & Pax, endemismes dels Alps i els Càrpats, respectivament. S’ha proposat que les tres espècies provenen d’un avantpassat comú (molt proper a D. elatum) que hauria donat lloc als tres taxons per fragmentació de l’àrea abans de la darrera glaciació (Pawlovsky, 1970). Totes aquestes espècies són tetraploides amb 2n = 32 cromosomes.
Sinònims del nom científic:
≡ Delphinium elatum subsp. montanum (DC.) Nyman
≡ Delphinium elatum var. montanum (DC.) Rouy & Fouc.
- Delphinium elatum auct.
- Delphinium pyrenaicum Pourr. (nom. inval.)
Distribució geogràfica
L’esperó de muntanya és un endemisme dels Pirineus orientals, principalment de la vora S. i E. de la fossa de la Cerdanya (serra del Cadí i massís del Puigmal), amb extensions fins al Pedraforca i el massís de Madres, cobrint una àrea de 60 km de llargada i uns 15 d’amplada, en territori administrativament espanyol i francès. Es coneix de nou localitats (tres de no retrobades recentment). A la Catalunya autònoma té presència confirmada actual en tres localitats: dues de la serra del Cadí (capçalera de la vall de Bastanist (Cerdanya) i serra Pedregosa (Berguedà) i una del massís del Puigmal (Coma de l’Orri, vall de Setcases, Ripollès). S’havia observat en una altra localitat del Cadí (Cava, Alt Urgell) no retrobada des del 1985, però insuficientment prospectada i al Pedraforca (Berguedà), població no retrobada des de 1984 i probablement extingida. A la Catalunya del Nord es troba en dues localitats a la serra del Puigmal (vall d’Eina i Cambra d’Ase), una al massís de Madres (Noedes) i una al coll de la Portella de Mantet, no retrobada des de 1896. Les indicacions antigues del massís de Sant Llorenç del Munt, provinents d’una citació de Cadevall de principis del s. XX, són errònies i corresponen en realitat a D. bolosii (Blanché, 1991).
|