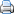|
FITXES ANTERIORS DE "La Planta del Mes" del Portal BioC
Sergi Massó
Silene sennenii Pau in Bol. Soc. Aragonesa Ci. Nat. 4: 309 (1905)
Silene sennenii és un colitx dedicat a Frère Sennen, botànic occità i germà de La Salle que treballà en la flora empordanesa a principis del S. XX. Es tracta d’una espècie endèmica de la comarca de l’Alt Empordà fortament amenaçada (categoria EN, “En perill”), que ha estat estudiada per investigadors del BioC des de fa anys i, actualment, és objecte del projecte de recerca CGL2007-60475/BOS. Ha estat part de la tesi doctoral de M.C. Martinell, del màster en ciències farmacèutiques de S. Massó i del treball dirigit de L. Guasch. Podeu veure el reportatge de TV3 sobre aquesta espècie emès l’any 2008, fent clic aquí.
DESCRIPCIÓ I IDENTITAT TAXONÒMICA
Silene sennenii és un hemicriptòfit amb tiges floríferes ascendents d’entre 15 i 80 cm (Figura 1A). Fulles estretament elíptiques a lanceolades, agudes, atenuades en el pecíol, amb pèls no glandulars. Flors actinomorfes, pentàmeres, proteràndriques i agrupades en dicasis, amb la corol·La de color blanc o rosat on els pètals, que mesuren entre 7 i 9 mm, es repleguen en les hores de insolació (Figura 1B). Pedicels de 4 a 8 mm, glabres. Bràctees més curtes que els pedicels, ovadolanceolades. Calze d’entre 13 i 15 mm, amplament ovat o suborbicular, amb el marge escariós i ample. Càpsules 8-9 x 4-5 mm amb nombroses llavors (40-70 per fruit) de 0,9-1 x 1-1,3 mm.

Figura 1. Individu desenterrat de Silene sennenii (A) i detall de les flors a primera hora del matí (B). [Fotografies: (A), M.C. Martinell; (B), S. Massó]
Nombre cromosòmic
És una espècie diploide de 2n = 24 (López-Pujol et al., 2007).
Taxonomia i nomenclatura
Silene sennenii Pau in Bol. Soc. Aragonesa Ci. Nat. 4: 309 (1905)
≡S. italica subsp. sennenii (Pau) O. Bolòs & Vigo in Butll. Inst. Cat. Hist. Nat. 38:87 (1974)
≡ S. saxíIraga subsp. sennenii (Pau) Malag. in Acta Phytoyax. Barcin. 18:9 (1976)
L’espècie va ser descrita pel botànic i farmacèutic de Sogorb Carlos Pau l’any 1905 i avui dia ofereix pocs dubtes respecte al seu rang específic (Chater & Walters, 1964; Jeanmonod, 1984; Talavera, 1990). Tot i això, existeixen dues flores (Malagarriga, 1976; Bolòs & Vigo, 1984) que la consideren a nivell de subespècie. Jeanmonod (1984) rebutja les combinacions de Bolòs & Vigo i de Malagarriga i referma la idea que es tracta d’una espècie ben diferenciada.
HÀBITAT
Silene sennenii creix gairebé exclusivament en fenassars (Brachypodietum phoenicoidis Br.-Bl., Figura 2), comunitat que, en la majoria de casos, està associada actualment a l’Alt Empordà als marges dels camps de cultiu, talussos i, excepcionalment, en cultius abandonats, en una alçada d’entre 15 i 100 metres. El comportament fitocenològic de la silene de Sennen va ser estudiat per Font Garcia & Gesti (2002). L’espècie pot aparèixer en altres ambients de manera secundària, tolerant una lleugera ruderalització del sòl. A part de Brachypodium phoenicoides, es troba habitualment acompanyada d’Eryngium campestre, Dactylis glomerata subsp. hispanica, Foeniculum vulgare, Ononis tridentata, Clematis flammula, Rubus ulmifolius, Silene vulgaris i Avena barbata.

Figura 2. Fenassar al marge d’un camp a la població del Riu Manol (RMA). [Fotografia: M.C. Martinell]
DISTRIBUCIÓ GEOGRÀFICA I NOMBRE D'EFECTIUS
Silene sennenii és un endemisme estricte de la comarca de l'Alt Empordà (Figura 3).
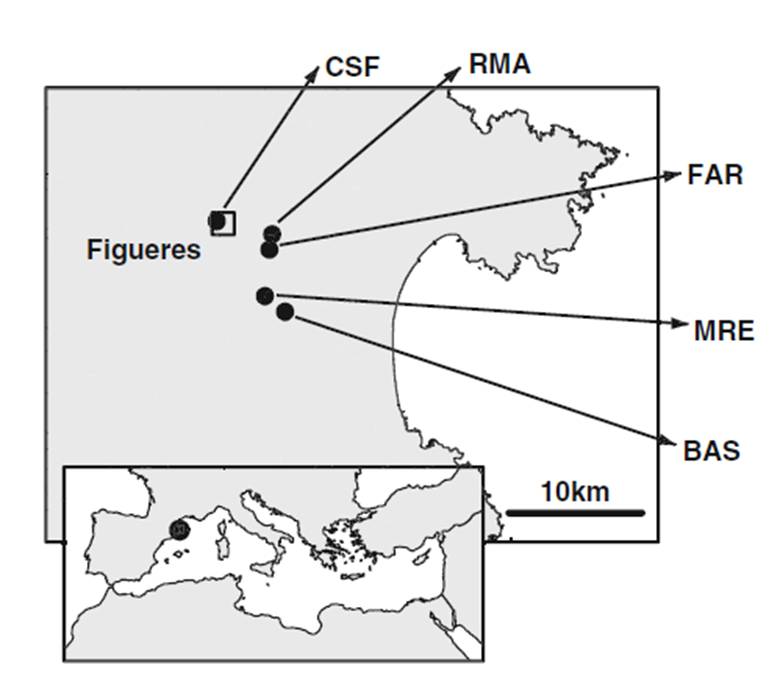
Figura 3. Àrea de distribució mundial de la silene de Sennen
Actualment es coneix de cinc localitats, distribuïdes en 13 quadrats UTM de 1 x 1 km de costat, a les rodalies de Figueres, en una de les quals (Castell de Sant Ferran, CSF) s’hi concentra més del 70% del total dels individus. Algunes poblacions del sector oriental de la seva àrea de distribució (Riumors, Fortià, Fortianell) semblen haver desaparegut (López-Pujol et al., 2007). El darrer cens complert data de 2008 (Martinell, 2010) i es van comptabilitzar un total de 4.308 individus. Les cinc localitats (Figura 4) són les següents:
- Castell de Sant Ferran (CSF) (31TDG97, DG98): Situada al voltant d’aquesta fortalesa militar del segle XVIII a les afores de Figueres, és la població més nombrosa -3097 individus l’estiu del 2008 (Martinell, 2010)-. Silene sennenii es troba als talussos
 artificials que envolten la fortalesa, saltant sovint als marges dels camps que l’envolten. Dins del castell es troba un petit nucli d’un centenar d’individus. artificials que envolten la fortalesa, saltant sovint als marges dels camps que l’envolten. Dins del castell es troba un petit nucli d’un centenar d’individus.
- Baseia (BAS) (31TEG07): És la segona població en nombre d’individus -1011 l’any 2008 (Martinell, 2010)-. Es troba dividida en
 cinc nuclis, dos dels quals força amplis, en prats no cultivats, marges de camps, i al llarg de pistes forestals i d’una carretera. cinc nuclis, dos dels quals força amplis, en prats no cultivats, marges de camps, i al llarg de pistes forestals i d’una carretera.
- Riu Manol (RMA) (31TDG97): Població lineal situada en un fenassar limitat per un camí a la riba esquerra del riu Manol, a l’alçada de
 Vilatenim. L’any 2008 aquesta població tenia 130 individus (Martinell, 2010). Vilatenim. L’any 2008 aquesta població tenia 130 individus (Martinell, 2010).
- El Far d’Empordà (FEM) 31TDG97: Població situada en un ampli talús entre un camp de conreu i el nucli urbà. Entre aquesta població i l’anterior (RMA) es troben alguns individus aïllats en marges de camins rurals i camps de cultiu. La població comptava amb 69 individus l’any 2008 (Martinell, 2010).
- Mas Renart (MRE) (31TDG97: Població que al 2008 comptava únicament amb un únic individu (Martinell, 2010).
Figura 4. Mapa de les localitats de Silene sennenii [Cartografia de base: ICC]
BIOLOGIA REPRODUCTIVA
Desconeguda fins fa ben poc, el projecte de recerca del BioC GL2007-60475/BOS ha aportat les primeres dades que caracteritzen la biologia reproductiva de la silene de Sennen, amb la col·laboració de diversos investigadors i d’estudiants en formació (Figura 5)

Figura 5. Mane Martinell i Laia Guasch, durant els treballs de biologia de la pol·linització de Silene sennenii, als fossats del Castell de Sant Ferran de Figueres (Alt Empordà). Agost de 2007. (Fotografia: L. Guasch).
Silene sennenii presenta pol·linització entomòfila, principalment per lepidòpters d’hàbits nocturns o crepusculars. Aproximadament un mes després de la pol·linització el fruit ja és madur.
Desenvolupament floral
El període de floració s’estén des del mes de juliol fins al mes d’octubre, amb especial intensitat a finals d’estiu. Cada tija (vara) produeix nombroses flors cada any. El seguiment fotogràfic del desenvolupament de cada flor al camp (Figures 6a i 6b) mostra els diversos estadis que es succeeixen al llarg de les tres o quatre nits d’obertura de cadascuna de les flors.
Es tracta d’una espècie proteràndrica: els botons florals madurs s’obren al capvespre, es despleguen els pètals i resten les flors obertes amb un primer verticil de 5 estams erectes, que contenen pol·len, fins al matí següent, en què els pètals es repleguen i la flor resta aparentment marcida (Figura 6 a). Al capvespre de la segona nit, novament s’obren les flors, que presenten un segon verticil de 5 estams dreçats i exposats, mentre que els 5 corresponents a la nit anterior apareixen vinclats i buits de pol·len, amb les anteres fora de l’abast dels pol·linitzadors i, passada la matinada, el matí següent tornen a replegar-se els pètals. Finalment, la tercera nit, el desplegament dels pètals deixa pas a l’estat femení de la flor, en aquest cas amb l’emergència de tres estils exerts que es mantenen oberts i receptius fins al matí següent, quan la flor torna a l’estat de repòs; cada flor pot romandre en aquesta fase femenina entre una i tres nits (Martinell et al., 2010) (Figura 6 b)

Figura 6a. Desenvolupament floral de Silene sennenii durant la primera nit (Fotos M.C. Martinell)

Figura 6b. Desenvolupament floral de Silene sennenii durant les segona i tercera nits (Fotos M.C. Martinell)
Censos de pol·linitzadors
Les observacions dels pol·linitzadors s’han dut a terme a intervals de 15 min al llarg de les 24 h del dia, durant la nit mirant de no pertorbar lumínicament les poblacions (a les fosques o, amb llum vermell si era imprescindible durant uns instants –vegeu vídeo).
Les diverses espècies d’insectes que visiten les flors de S. sennenii són principalment lepidòpters d’hàbits nocturns o crepusculars (Rhodometra sacraria, Antigastra catalaunalis, Calophasia platyptera, Macdunnoughia confusa, Autographa gamma, Mythimna farrago, Herse convolvuli i Macroglossum stellatarum), encara que també s’han observat casos aïllats de visites diürnes per part d’himenòpters (Lassioglossum sp., Amegilla sp. i Apis mellifera) (Martinell et al., 2010). El nombre de visites al llarg de 24 h mostra una corba amb un màxim durant les hores crepusculars i de l’alba i un mínim a les hores centrals del dia (Figura 7).
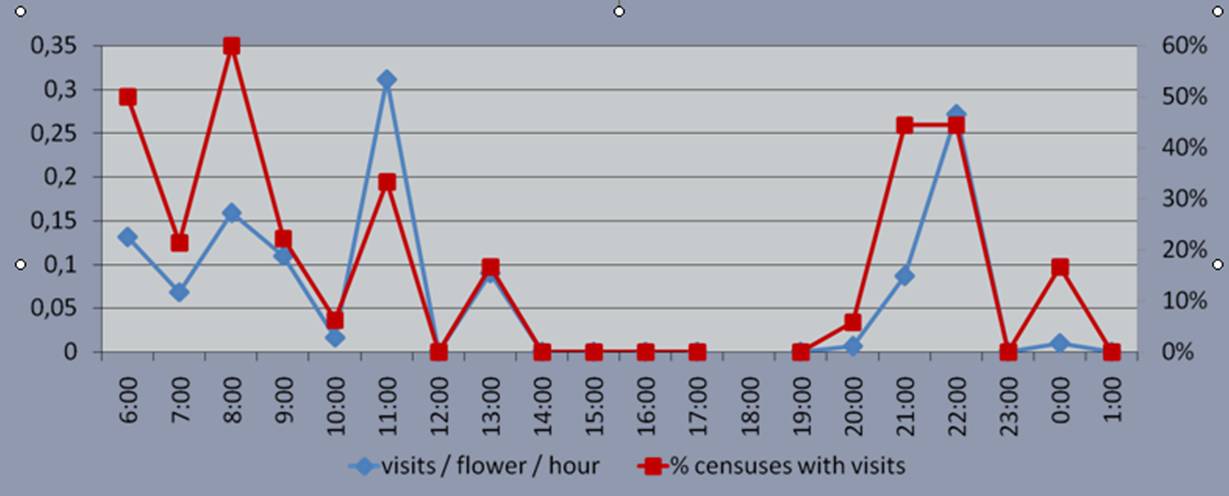
Figura 7. Corba de nombre de visites de pol·linitzadors a Silene sennenii (nombre de visites per hora [blau] i nombre de censos amb visites [vermell])
Existeix, doncs, un acoblament molt acurat entre el ritme d’obertura de les flors i el de visites de pol·linitzadors durant les nits, així com entre les morfologies floral (peces florals de color clar –rosat a blanc- que defineixen un tub d’accés relativament estret) i de la probòscide els principals lepidòpters visitants nocturns (llargues espiritrompes que poden atènyer el nèctar produït al fons de la flor), amb efectivitat comprovada per les càrregues pol·líniques dipositades al cos dels animals. La sincronització, però, no és perfecta del tot i durant les hores diürnes, malgrat les flors tancades i menys vistoses, també es produeixen visites (Figura 7), en aquest cas principalment d’un halíctid del gènere Lasioglossum, que també transporta pol·len.
Experiments d’exclusió
A través d’embossaments selectius de flors per restringir l’accés d’insectes visitants de manera voluntària es pot avaluar l’aportació de pol·len en cada període de temps (per recompte dels grans de pol·len dipositats als estigmes) i l’efectivitat final (estimada com a nombre de les granes produïdes en cada condició).
Aquests experiments mostren coincidència en els resultats les càrregues pol·líniques als estigmes, el nombre de flors que fan fruit i el nombre de llavors per fruit (Figura 8) i permeten concloure que:
1. Malgrat la proteràndria ben marcada, S. sennenii és capaç d’autopol·linitzar-se espontàniament, encara que amb producció de granes significativament reduïda.
2. Les flors pol·linitzades de nit esdevenen fruits en una proporció més gran i fan més llavors que les pol·lnitzades de dia o les flors tancades i, per tant, encara que hi hagi menys visites totals durant la nit, els pol·linitzadors nocturns són molt més eficients que no pas els diürns, que també n’hi ha.
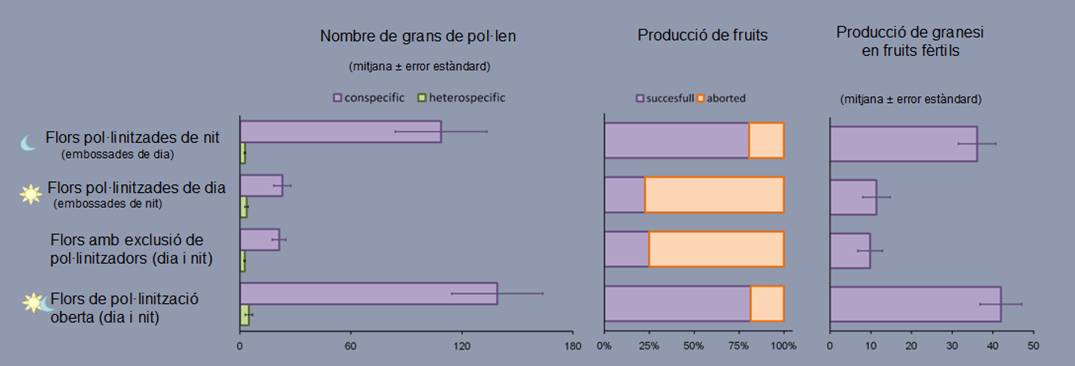
Figura 8. Resultats dels experiments d’exclusió d’insectes en Silene sennenii
Els motius de la manca d’eficiència dels pol·linitzadors diürns no són ben coneguts del tot. D’una banda, llur comportament durant les visites pot ser poc eficient per tal com acumularien el pol·len en parts del cos en què el contacte amb els estigmes no és possible, però, d’altra banda, podria ser que la qualitat del pol·len disponible i/o de la receptivitat de l’estigma durant el dia es trobés disminuïda, prevenint així serveis de pol·linització poc adequats. Massó (2011) ha comprovat aquesta darrera hipòtesi, que es confirma atès que, després del seguiment hora a hora de l’estat de maduració dels grans de pol·len, el punt àlgid (quan tenen més probabilitat de germinar i formar els tubs pol·llínics) el tindrien al voltant de la mitjanit. És precisament en aquest moment quan els potencials pol·linitzadors de S. sennenii visiten les flors al camp (Martinell, 2010). A mesura que avança la nit, la viabilitat del pol·len va disminuint i el pol·len que ja no és viable es desprèn de les anteres, que a les hores diürnes només conserven pocs pòl·lens i, encara, amb escassa capacitat germinativa. Així doncs, existeix una bona sincronització entre la presentació de recompenses, el desenvolupament dels òrgans florals, la fisiologia del desenvolupament del pol·llen i l’activitat dels pol·linitzadors per a un òptim nocturn.
Emissió d’aromes
Finalment, la síndrome de pol·linització nocturna es pot acabar de caracteritzar per l’acoblament de l’emissió d’aromes, que actuen com a senyals d’atracció que guien les aproximacions dels pol·linitzadors, amb la resta de ritmes sincronitzats que acabem de descriure. S’han pres mostres d’aromes de flors individuals a diversos intervals de temps. Les mostres han estat analitzades per cromatografia de gasos acoblada a espectrometria de masses en col·laboració amb l’equip del Prof. S. Dötterl de la Universitat de Bayreuth, a Alemanya (Martinell et al., 2010)
S’ha pogut comprovar que Silene sennenii emet grans quantitats d’aromes durant la nit, emissió que es veu molt reduïda durant el dia. Les anàlisis han revelat que el constituent principal de l’aroma és el (E)-ß-ocimè. Aquest monoterpè (requadre a la Figura 9) seria el responsable de l’atracció dels pol·linitzadors, com s’ha demostrat en d’altres espècies de Silene i d’altres gèneres (Martinell et al., 2010).
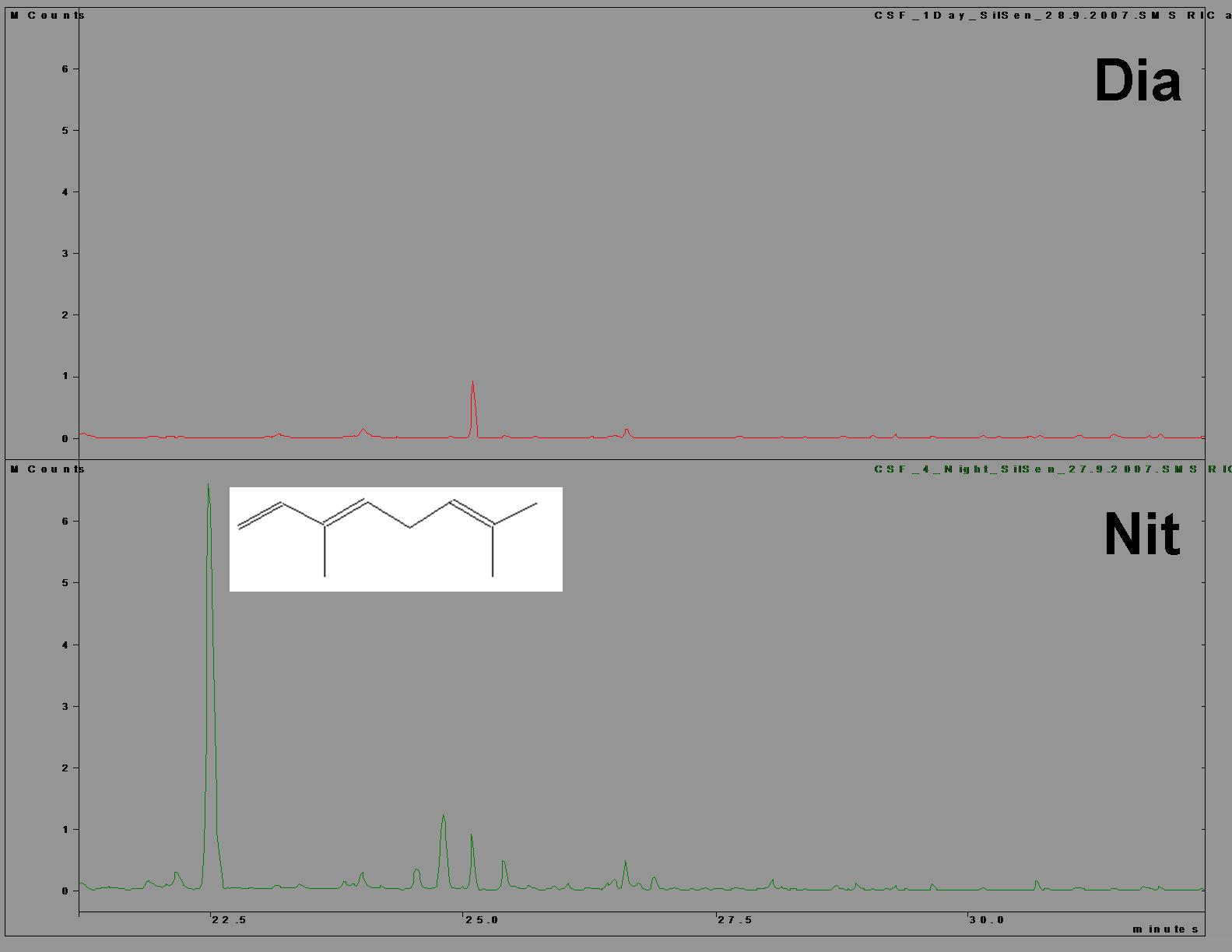
Figura 9. Cromatograma (GC-MS) d’una mostra d’aroma recollida durant el dia (a) i durant la nit (b).
Germinació
Les granes produïdes van madurant fins a finals d’estiu, quan atenyen l’estat de maduresa i la capacitat de contribuir a la generació següent. Durant el decurs del projecte s’ha avaluat diverses vegades la capacitat germinativa de les llavors, entre d’altres motius com a guia per a la conservació ex situ de les granes preservades en bancs de germoplasma i per al coneixement de possibles limitacions en cadascuna de les fenofases del cicle de l’espècie.
La darrera avaluació (Massó, 2011) confirma que la germinació de les granes no és un factor limitant de rellevància. Així, de 215 llavors sembrades, 187 (86,98% del total) havien germinat al cap de 5 dies (Figura 10)
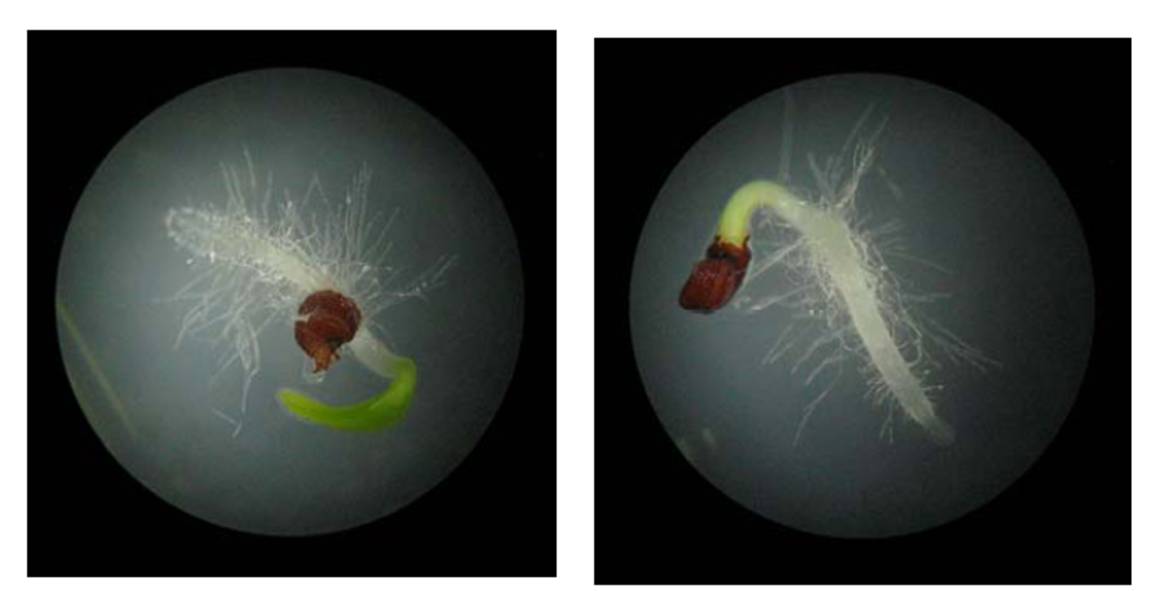
Figura 10. Dues llavors de S. sennenii als 2 dies d’haver germinat. [Fotografies S. Massó]
Pel que fa a la variabilitat interpoblacional de la capacitat germinativa, s’ha detectat que existeixen diferències significatives únicament en la població del Riu Manol (RMA) (Xi quadrat, X =6,80, p = 0,0333), on el percentatge de germinació és menor, fet que coincideix amb una mida poblacional més petita, uns serveis de pol·lnització de menys qualitat, menor diversitat genètica i, en general un hàbitat més alterat i amb efectes evidents de fragmentació (vegeu més endavant). També s’ha comprovat que les llavors de la silene de Sennen triguen més en germinar a mesura que envelleixen: això suggereix, d’una banda, que no és esperable una estratègia de banc de llavors al sòl de manera permanent i, de l’altra, que la preservació en condicions adequades als bancs de germoplasma és necessària si les llavors es volen conservar a llarg termini.
Establiment de plàntules i supervivència
Les llavors germinades han estat transplantades a testos i safates al Servei d’Hivernacles i Vivers de la Facultat de Farmàcia. Malgrat una certa mortalitat de les plàntules, les taxes de supervivència del primer any són molt satisfactòries, amb valors molt similars en rèpliques de dos anys seguits: 80,6% (Martinell, 2010), 80,5% (Massó, 2011) realitzades en condicions experimentals equivalents. Com a resultat, l’equip ha establert una metodologia estandarditzada de cultiu ex situ.

Figura 11. Safates amb individus de Silene sennenii provinents de llavors cultivats al SHV de la Facultat de Farmàcia la tardor de 2010 [Foto: S. Massó]
El pas de fenofase juvenil cap a reproductor presenta nivells d’incertesa més grans. Les dades obtingudes indiquen que un 72,55% van sortir de l’estat vegetatiu, formant almenys una vara florífera. Tanmateix, tan sols un 18,3% van produir flors el 2010 (Massó, 2011), valor molt allunyat dels trobats per Martinell (2010) en les mateixes èpoques de l’any: 70,6% i 80,6% de floració en 2008 i 2009 respectivament. Aquesta variabilitat interanual està relacionada amb les condicions meteorològiques i pot significar també un important retard fenològic d’entre 5 i 8 setmanes segons l’any, tant en el desenvolupament de l’individu com en el període de floració. Les corbes de creixement revelen que S. sennenii necessitaria una temperatura superior als 20ºC per tal d’iniciar el cicle de floració i que si se sobrepassa una determinada temperatura màxima, l’individu s’asseca, amb risc de mortalitat malgrat l’aparició de rebrots tardorals ocasionals. La recomanació per al cultiu ex-situ és de transplantament a recipients més grans el segon any i de vetllar pel manteniment dels nivells d’humitat.
DIVERSITAT GENÈTICA
S’ha estudiat la diversitat genètica d’aquesta espècie mitjançant electroforesi d’al·loenzims i en l’actualitat es duen a terme assaigs amb marcadors de més resolució (cpDNA, AFLPs, microsatèl·lits) als laboratoris de l’Institut Botànic de Barcelona, en col·laboració amb Sara López Vinyallonga.
Els resultats de l’anàlisi isoenzimàtica de les cinc poblacions romanents (López-Pujol et al., 2007), a partir de 21 loci interpretables, mostren que la diversitat genètica mitjana de la silene de Sennen és baixa (P = 20.9%, A = 1.31 i He = 0.071, Taula 1), força per sota d’altres espècies de Silene comunes i d’àrea extensa.
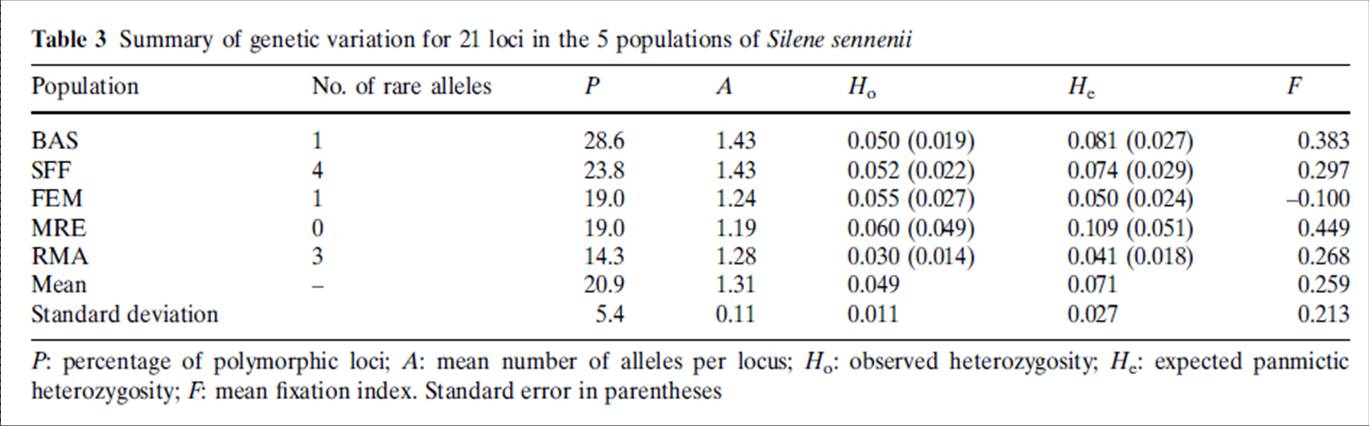
Taula 1. Paràmetres de diversitat isoenzimàtica a les poblacions de Silene sennenii (Font: López-Pujol et al. 2007)
A part de la població MRE, massa petita per considerar-ne els valors com a significatius, les dues poblacions més nombroses (CSF i BAS) presenten nivells de diversitat més elevats que les poblacions més petites, per a tots els paràmetres (Taula 1). L’erosió genètica deguda a la mida poblacional reduïda es revela pel nombre d’al·lels. En total es van detectar tan sols 30 al·lels. En les dues poblacions més grans (CSF i BAS) es van detectar tots 30 al·lels, mentre que a RMA i FEM se’n van detectar 27 i 26 respectivament. Per la seva part, la població més petita (MRE) mostrava únicament 25 al·llels.
Els valors d’heterozigosi observada (Ho) són en general més baixos que els d’heterozigosi esperada (He) i es detecta dèficit d’heterozigots a tots els loci polimòrfics de totes les poblacions (López-Pujol et al. 2007). L’excés d’homozigots també es manifesta en els valors de F i els quocients de consanguinitat de Wright (Fis = 0.25).
La divergència genètica entre poblacions, estimada pel paràmetre Fst, que mesura la diferenciació interpoblacional, dóna un valor elevat (Fst=0,27), indicatiu que una fracció significativa de la variabilitat genètica de S. sennenii és atribuïble a diferències entre poblacions (fet que caldrà tenir en compte en les mesures de gestió i conservació que s’emprenguin). De tota manera, diverses anàlisis de les matrius de diferenciació genètica i les distàncies geogràfiques no donen suport a un model d’aïllament per distància, que tampoc apareixen al dendrograma UPGMA (Figura 12): sense anar més lluny, les poblacions més semblants (CSF/SFF i BAS) són les geogràficament més allunyades (vegeu Figura 4).
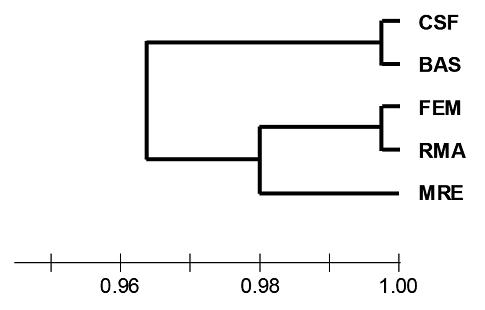
Figura 12. Dendrograma UPGMA de distàncies genètiques de Nei per a les poblacions de Silene sennenii (Font: López-Pujol et al., 2007)
La fragmentació o destrucció dels fenassars ocupats per la silene de Sennen pot explicar, almenys en part, els baixos nivells de diversitat genètica detectats com a causa de la reducció de mida poblacional i de l’aïllament entre els fragments romanents.
La bona correlació entre nombre d’al·lels i mida poblacional pot ser indicativa de deriva genètica a les poblacions, atès que els efectes de la deriva són més grans a les poblacions petites i, efectivament, els al·lels que es troben en proporcions més baixes a les poblacions grans (CSF i BAS) s’haurien perdut a les més petites (FEM i RMA).
L’estructura en rodals de les poblacions (que afavoreix encreuaments entre individus propers) accentua l’efecte dels baixos nivells de diversitat, amb dèficit d’heterozigots per autogàmia biparental (encreuament entre individus genèticament molt propers) en combinació amb l’efecte de l’autopol·linització que pot existir sota règims de pol·linització pertorbats.
Tot i que les cinc poblacions romanents són separades per pocs quilòmetres, els valors de divergència (Fst mitjana= 0,27) es troben per sobre dels reportats per a tàxons subjectes a fragmentació (10-20 %, López-Pujol et al, 2007), cosa que indicaria que la fragmentació està tenint un impacte en la composició genètica de les poblacions.
La desaparició d’algunes poblacions corresponents a citacions antigues o a nuclis intermedis entre les poblacions actuals, la reducció de mida de les romanents (per exemple, FEM es va reduir de 295 a 49 individus el 2001, a conseqüència de l’ampliació d’una carretera) amb els colls d’ampolla genètics subsegüents o la pressumpta fundació de nous nuclis a partir de pocs individus durant la construcció del castell de Sant Ferran (CSF) al s. XVIII són altres factors que haurien influït en el patró de diferenciació genètica detectat en l’actualitat.
CONSERVACIÓ
Amenaces
1-La principal amenaça a la qual està sotmesa aquesta espècie és la disminució de superfície i la pèrdua de qualitat del seu hàbitat per l’expansió de zones urbanitzades i les seves conseqüències (López-Pujol et al., 2007; Martinell, 2010) i per la transformació en cultius de regadiu d’alguns conreus al voltant dels quals creix (J. Font, com. pers.). Aquesta disminució i pèrdua de qualitat de l’hàbitat comporta una disminució de la població, més ràpida als nuclis més petits, tal com s’ha pogut comprovar per l’evolució de la demografia (Molero, 1987; Font Garcia et al., 2003; López-Pujol et al., 2007; Taula 2), en algun cas al límit de l’extinció (MRE: 4 individus el 2001; 1 únic individu el 2008).
|
Població
|
Cens 2001 (1)
|
Cens 2008 (2)
|
Variació
|
|
CSF
|
3209
|
3097
|
- 3,49%
|
|
BAS
|
1084
|
1011
|
-6,73%
|
|
FEM
|
49
|
69
|
+ 40,81%
|
|
RMA
|
172
|
130
|
-24,41%
|
|
MRE
|
4
|
1
|
-75%
|
|
TOTAL
|
4518
|
4308
|
-4,64
|
Taula 2. Censos poblacionals de la silene de Sennen corresponents als anys 2001 i 2008. Fonts: (1) López-Pujol et al., 2007; (2) Martinell (2010). Observacions: a) a (2) s’han inclòs individus de l’interior del recinte del castell que no havien estat detectats en anys anteriors; b) Les poblacions BAS i RMA van sofrir alteracions significatives amb posterioritat al cens, que van suposar la pèrdua d’efectius (cf. Notícia al Portal Bioc de 25/11/2009)
La mida extremadament reduïda de tres de les poblacions (de l’ordre d’un centenar o menys d’individus), les deixa al llindar o per sota de la mida poblacional mínima (MVP), incapaces de fer front a esdeveniments ocasionals catastròfics de caràcter estocàstic (fonamentalment per intervencions humanes), però també a limitacions de caràcter genètic, com reflecteixen les dades isoenzimàtiques que detecten elevades taxes d’endogàmia o de consanguinitat. Es desconeix l’àrea (i fins i tot l’hàbitat) que l’espècie devia ocupar en una comarca poc alterada fa segles, però l’assumpció actual és que és restringida a fenassars marginals (Font Garcia & Gesti, 2002), l’àrea dels quals es va reduint en quantitat i qualitat a gran velocitat, de manera que llur preservació és essencial per a la conservació de la silene de Sennen.
2-Al damunt d’aquesta situació s’afegeixen afectacions d’ importància diversa a conseqüència d’intervencions sobre les poblacions en els darrers cinc anys (vegeu-ne reportatge al Portal BioC ):
- estassades a les quals es sotmeten els marges de camins o carreteres a finals d’estiu que afecten de forma directa el cicle reproductiu de la planta, ja que en l’època en què es fan no tots els fruits han madurat i no s’ha acabat de produir la dispersió de les granes.
- arranjament de camins, explanacions i remoció de sòls per a la instal·lació d’un parc de plaques solars a la població BAS, que han soterrat c. 10% dels individus preexistents.
-alteració del fenassar i arrencament de diversos individus a conseqüència de la renovació i manteniment dels suports de transport elèctric (RMA)
I, en general, les conseqüències derivades de la freqüentació, abocament de brossa, incendis ocasionals, etc. a causa de la forta pressió humana sobre els retalls d’hàbitat romanent ocupat per l’espècie.
3-També es troba amenaçada de forma important la xarxa de pol·linitzadors (Martinell et al., 2010). D’una banda, la pol·linització nocturna pot veure afectada la qualitat i la quantitat dels serveis dels pol·linitzadors si es produeix una modificació de les condicions ambientals durant la nit. La població RMA, per exemple, té un nombre més elevat de visites per pol·linitzadors diürns que la resta, fet que pot interpretar-se perquè es troba en una zona il·lluminada artificialment i on no són tant freqüents els pol·linitzadors nocturns de S. sennenii. El resultat atribuible a aquest fet és una menor producció de llavors. La il·luminació dels entorns del Castell de Sant Ferran (CSF) prevista en projectes d’arranjament futur haurà de tenir en compte aquest factor.
D’altra banda, la qualitat general de l’hàbitat i la mida dels fragments romanents es troba en relació directa amb les taxes de visites d’insectes legítims i d’èxit reproductiu que, a les poblacions més petites mostra símptomes de disrupció del sistema de mutualisme planta-pol·linitzador, amb conseqüències sobre el manteniment de la diversitat genètica i la qualitat dels nous reclutaments.
4-Algunes espècies de Silene tenen reportades infeccions fúngiques i predació per agents diversos (p.e. Microbotryum), en règims gairebé mutualístics, molt difícils d’erradicar. L’anàlisi duta a terme per especialistes (cf. Martinell, 2010), indica que la silene de Sennen és lliure d’aquestes infeccions en l’actualitat.
Avaluació del grau d’amenaça segons criteris UICN
Categoria UICN per a Catalunya: EN B1ab(i,ii,iii,iv,v)+2ab(i,ii,iii,iv,v)
Categoria UICN mundial: íd.
Font: Sàez et al., 2010 (segons criteris UICN, 2001)
Avaluacions anteriors
R (“rara”, categoria UICN 1984) a Molero, 1987
EN (“en perill”) B1ab(i,ii,iii,iv,v)+2ab(i,ii,iii,iv,v) a Font Garcia et al., 2003
EN (“en perill”) B1ab(i,ii,iii,iv,v)+2ab(i,ii,iii,iv,v) a Font Garcia et al., 2006
Protecció legal
L’espècie és catalogada com a “en perill d’extinció” en l'Annex I del Catàleg de flora amenaçada de Catalunya (DOGC, 2008). No existeix cap mena de protecció legal a nivell estatal o europeu per a Silene sennenii.
Mesures de conservació en curs
En l’actualitat s’estan aplicant les següents mesures de conservació i recuperació (segons Martinell, 2010 i dades inèdites):
- Assaigs de germinació i cultiu al Jardí Botànic Marimurtra de Blanes (Fundació Carl Faust), a través de la inclusió d’aquesta espècie dins el projecte Phoenix2014, de la Xarxa Iberomacaronèsica de Jardins Botànics. Així mateix, s’hi estan duent a terme la posada a punt de protocols de germinació i cultiu.
- Conservació de 16.750 llavors al banc de germoplasma del Jardí Botànic Marimurtra de Blanes (Fundació Carl Faust). Les llavors provenen de CSF (9.000), BAS (5.800), RMA (1.550) i FEM (400) i van ser totes collides l’any 2008 en el marc de l’ Earthwatch Project, amb la col·laboració del BioC.
- Cultiu de 43 individus al Jardí Botànic Marimurtra de Blanes (Fundació Carl Faust) i d’uns 250 al viver de la Facultat de Farmàcia de la Universitat de Barcelona. Aquests darrers han estat emprats per a la realització d’aquest estudi i, havent finalitzat recentment, han estat traslladats al Jardí Botànic de Barcelona (vegeu Notícia 1/6/2011 al Portal BioC)
- Conservació de l’hàbitat al Castell de Sant Ferran (CSF) atès l’estatus militar original del recinte, que impedeix la construcció i la intervenció en els talussos que l’envolten.
- Conservació preventiva de l’hàbitat a càrrec de la Generalitat de Catalunya, a través dels serveis territorials a Girona del Departament d’Agricultura, Ramaderia, Pesca, Alimentació i Medi Natural, mitjançant la informació de l’existència d’aquesta espècie protegida a ajuntaments, particulars i empreses propietàries o gestores dels terrenys que ocupa.
- Inclusió dels perímetres dels polígons ocupats per l’espècie al Mapa d’Àrees d’interès per a la flora i la fauna del Departament d’Agricultura, Ramaderia, Pesca, Alimentació i Medi Natural.
- Inici dels estudis de mesures de reforçament poblacional i de pla de recuperació per part del Departament d’Agricultura, Ramaderia, Pesca, Alimentació i Medi Natural.
Mesures de conservació proposades
Redacció d’un pla de recuperació específic urgent a causa de la vulnerabilitat de les poblacions que encara es conserven, tal com preveu el Decret 172/2008, que inclogui:
- Creació d’una reserva per a la conservació concertada (biodiversitat i monuments històrics) al castell de Sant Ferran (CSF), amb caràcter equivalent al de microrreserva, on es conserva la totalitat de la diversitat genètica coneguda de l’espècie (Blanché, 2008, López Pujol et al., 2007)
- Reforçament poblacional de les poblacions més petites (< 100 individus)
- Mesures de gestió dels hàbitats de fenassars romanents que incloguin la regulació dels usos agrícoles de les finques limítrofs
- Afavoriment de la connectivitat dels nuclis coneguts
- Estudi de la dinàmica poblacional de manera experimental (anàlisi de viabilitat poblacional, AVP) i de marcadors moleculars de més resolució
- Continuació de les tasques de conservació ex situ i estudi de protocols de germinació, aclimatació i transplantament
- Gestió adequada de la il·luminació a les àrees periurbanes que afectin les poblacions
- Gestió de l’hàbitat que afavoreixi la provisió de refugi i d’alimentació de les larves dels pol·linitzadors de S. sennenii.
Legals: Inclusió de l’espècie al Catálogo Español de Especies Amenazadas i al llistat d’espècies prioritàries de la Directiva Hàbitats amb la màxima categoria.
VEURE VÍDEO
"Silene sennenii, una empordanesa en perill". Emissió al programa “El Medi Ambient”, TV3, 25/11/2008
REFERÈNCIES BIBLIOGRÀFIQUES
Blanché, C., 2008. Conservació concertada de flora i de monuments històrics. E-Opinió, 2. BioC, Portal de Biologia de la Conservació de Plantes. [Publicat en línia 13.01.2008] http://bioc.org.es/bioc/index.php?option=com_content&view=article&id=57:flora-i-monuments&catid=20:e-opini&Itemid=54
Bolòs, O. & Vigo, J., 1984. Flora dels Països Catalans. Barcino, Barcelona.
Chater, O. & Walters, S.M., 1964. Silene L. In: Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M. & Webb, D.A. (eds.). Flora Europaea. Vol. 1. Cambridge University Press, Cambridge, pp. 158-181.
DOGC, 2008. Decret 172/2008, de 26 d’agost, de creació del Catàleg de flora amenaçada de Catalunya. Diari Oficial de la Generalitat de Catalunya 5204: 65881-65895.
Font Garcia, J. & Gesti, J., 2002. Precisiones corológicas y comportamiento fitosociológico del endemismo Silene sennenii Pau. 1er Congreso de Biologia de la Conservación de Plantas. València, 2-5 octubre de 2002.
Font Garcia, J., Gesti, J. & Sàez, L., 2003. Silene sennenii Pau In: A. Bañares, G. Blanca, J. Güemes, J.C. Moreno & S. Ortiz (eds). Atlas y Libro Rojo de la flora vascular amenazada de España. Dirección General de Conservación de la Naturaleza, Madrid, pp. 858-859.
Font Garcia, J., Gesti , J. & Sáez, L. 2006. Silene sennenii In: IUCN 2010. IUCN Red List of Threatened Species. Version 2010.4. <www.iucnredlist.org>. Downloaded on 20 May 2011.
Gesti, J., 2006. El Poblament vegetal dels Aiguamolls de l’Empordà. Institut d’Estudis Catalans, Secció de Ciències Biològiques. Arxius de les Seccions de Ciències, 138. Barcelona.
Hamrick, J.L. & Godt, M.J.W., 1990. Allozyme diversity in plant species. In: Brown, A.H.D.; Clegg, M.T.; Kahler, A.L. & Weir, B.S. (eds.). Plant population genetics, breeding and genetic resources. Sinauer Associates, Sunderland (MA, USA): 43-63.
Jeanmonod, D., 1984. Révision de la section Siphonomorpha Otth du genre Silene L. (Caryophyllaceae) en Mediterranée occidentale. IV: Species caeterae. Candollea, 40: 5-34.
López-Pujol, J.; Font, J.; Simon, J. & Blanché, C., 2007. Can the preservation of historical relicts permit the conservation of the endangered plant species? The case of Silene sennenii (Caryophyllaceae). Conservation Genetics, 8: 903-912.
López-Pujol, J.; Bosch, M.; Simon, J. & Blanché, C., 2008. Diversitat isoenzimàtica de la flora vascular silvestre dels Països Catalans. Butlletí de la Institució Catalana d’Història Natural. 74: 5-28.
Malagarriga, R., 1976. Catálogo de las plantas superiores del Alt Empordà. Acta Phytotaxonomica Barcinonensia, 18: 1-146.
Martinell, M.C., 2010. Biología de la conservación de especies amenazadas de área de distribución restringida en Cataluña. Tesi doctoral. Universitat de Barcelona, Barcelona.
Martinell, M.C.; Dötterl, S.; Blanché, C.; Rovira, A.; Massó, S. & Bosch, M., 2010. Nocturnal pollination of the endemic Silene sennenii (Caryophyllaceae): an endangered mutualism? Plant Ecology, 211 (1): 203-218.
Massó, S., 2011. Estudis de Biologia de la reproducció i Conservació ex situ de l'endemisme empordanès Silene sennenii. Màster Experimental en Ciències Farmacèutiques (especialitat Botànica). Facultat de Farmàcia, Universitat de Barcelona (inèd).
Molero, J., 1987. Silene sennenii In: C. Gómez-Campo (ed.), Libro Rojo de Especies Vegetales Amenazadas de España Peninsular e Islas Baleares. ICONA, Madrid: 582-583
Pau, C., 1905. Plantes observées dans l’Ampourdan (surtout aux environs de Figueras) pendant l’année 1905 par le frère Sennen. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 4: 303-332.
Sáez, L.; Aymerich, P. & Blanché, C., 2010. Llibre vermell de les plantes vasculars endèmiques i amenaçades de Catalunya. Argania Edicions, Barcelona
Talavera, S., 1990. Silene L. In: Castroviejo, S. (Ed.). Flora iberica. Real Jardín Botánico de Madrid, Madrid, pp. 313-406.
UICN, 2001.Categorías y Criterios de la Lista Roja de la UICN: Versión 3.1. Comisión de Supervivencia de Especies de la UICN., Gland, & Cambridge.
CITACIÓ - HANDLE i Versió PDF
- Citació recomanada: Massó, S. 2011. Planta del Mes núm. 8: Silene de Sennen. Portal de Biologia de la Conservació de plantes. Laboratori de Botànica, Facultat de Farmàcia, Universitat de Barcelona. URI: http://hdl.handle.net/2445/32973
|